1- مقدمه
2- موتورهای مولکولی پروتئینی پایه ATP
2-2- موتورهای میوزین و کینسین
3- موتورهای مولکولی پایه DNA
4- موتورهای مولکولی غیرآلی (شیمیایی)
نتیجه گیری
1- مقدمه
انرژی مورد نیاز برای تغییرات ساختاری در موتورهای مولکولی، از منابع گوناگونی تأمین میشود. این انرژی بسته به نوع موتور مولکولی میتواند از جنس شیمیایی، نور و یا تغییرات محیطی مانند تغییر دما یا pH محیط باشد. در واقع، موتورهای مولکولی دارای ساختار فضایی مشخص با قواعد معینی هستند که تحریک خارجی باعث تغییر ساختار و قواعد بخشی از موتور میشود و نتیجه آن، به وجود آمدن حرکتهای مکانیکی است.
میلیونها سال است که در طبیعت، انواع مختلفی از موتورهای مولکولی وجود دارند که در گذر سالیان متمادی، عملکرد آنها بهینهسازی شده است. همانطور که شناخت ما نسبت به موتورهای مولکولی طبیعی در حال افزایش است، توانایی تقلید ما از آنها برای سنتز گونههای مصنوعی نیز در حال توسعه است. لذا میتوان موتورهای مولکولی را به دو گروه اصلی موتورهای طبیعی (پروتئینی) و موتورهای مصنوعی تقسیمبندی کرد. موتورهای طبیعی اغلب مبتنی بر ATP هستند. در مقاله حاضر به معرفی انواع مختلف موتورهای ملکولی و مکانیزم علمکرد آنها پرداخته خواهد شد.
2- موتورهای مولکولی پروتئینی پایه ATP
1-2- موتورهای F0-F1-ATPase
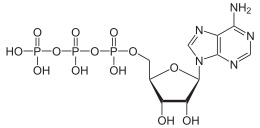
ATP واحد رایج انرژی در سیستمهای مولکولی زیستی است. این مولکول متشکل از 3 یون فسفات است که به پایه آنزیمی مولکول وصل هستند (شکل 1). با هیدرولیز ATP و شکسته شدن پیوند موجود بین «یونهای فسفات» و «پایه آنزیمی مولکول ATP» (یعنی ADP)، انرژی این ملکول آزاد میشود و میتواند فرآیندهای زیستی را به پیش ببرد. این واکنش در رابطه 1 خلاصه شده است:
(معادله 1) ATP+H2O←→ADP+Pi
همانطور که مشاهده میشود، شکسته شدن پیوند ATP (یعنی مصرف آن) باعث تولید ملکول ADP (adenosine diphosphate) و یون فسفات (Pi) میشود. ترکیب دوباره این دو محصول میتواند مولکول ATP را مجدداً تولید کند. بنابراین، معادله 1، یک واکنش برگشتپذیر است. بر خلاف فرآیند هیدرولیز ATP که باعث تولید انرژی میشود (یعنی رابطه 1 خود به خودی است)، ترکیب مجدد محصولات واکنش و تولید دوباره ATP نیاز به انرژی دارد. این واکنش، اساس عملکرد موتور مولکولی F0-F1-ATPase است.
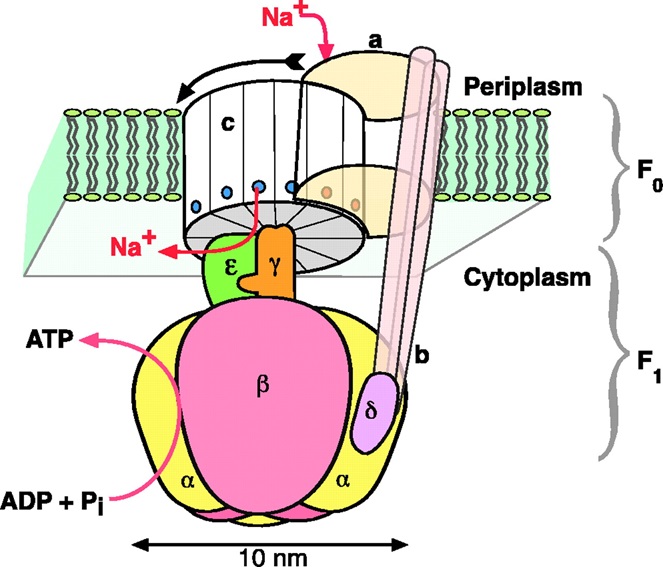
در واقع، موتور ملکولی F0-F1-ATPase، ترکیب دو موتور هیدروفوبیکی درونغشایی F0-ATPase (hydrophobic transmembrane) و موتور گلوبولار F1-ATPase است که ساختار و عملکردهای متفاوتی نسبت به یکدیگر دارند. موتور F0 ساختارهای فضایی متفاوتی در اندامهای مختلف دارد و نحوه عملکرد آن، به منبع انرژی درون آن اندام وابسته است. به عنوان مثال، در میتوکندری (mitochondria) سلولهای تشکیل دهنده بافت حیوانات (که اندامکی در داخل سلولها برای تبدیل مواد غذایی به ATP است)، موتور F0 در غشاء داخلی میتوکندری قرار گرفته است و نیروی محرکه یونی (ion-motive) باعث حرکت آن میشود. در واقع، اختلاف غلظت یون در دو طرف موتور باعث ایجاد یک شار یونی میشود و این شار، حرکت موتور را به وجود میآورد. اگرچه در ابتدا تصور میشد که نیروی محرکه این موتورها تنها «نیروی محرکه یونی حاصل از یون هیدروژن» است، اما امروزه مطالعات نشان داده است که یون سدیم نیز باعث حرکت آنها میشود. این نیروی محرکه یونی به صورت کار بر واحد یون تعریف میشود که یک یون میتواند با حرکت از بین غشا آن مقدار کار را تولید کند.
ساختار موتور F1: این موتور که انرژی خود را از هیدرولیز ATP به دست میآورد، از یک ستون پروتئینی مرکزی به نام «زیرواحد-γ» (γ-subunit) تشکیل شده است. این ستون توسط 6 زیرواحد دیگر به نامهای α و β محاصره شده است. زیرواحدهای α و β به طور یکدرمیان و به صورت متقارن در کنار یکدیگر قرار گرفتهاند. همچنین، زیرواحد δ به محیط استوانهای α- β متصل است و زیرواحد Ɛ در زیر یا کنار ستون γ قرار دارد. زیرواحدهای α و β حاوی محلهایی معینی برای اتصال آنزیمها هستند. این مکانها محل مناسبی برای اتصال مولکولهای ATP/ADP به شمار میروند. محلهای اتصال آنزیمها در زیرواحدهای α فقط به آنزیمها اجازه اتصال فیزیکی میدهد، درحالی که محلهای اتصال در زیرواحدهای β، نقش کاتالیزور واکنش (تبدیل ATP به ADP یا بالعکس) را دارند. شکل 2 شمایی از ساختار موتور F1 را نشان میدهد که بخشی از یک باکتری است.
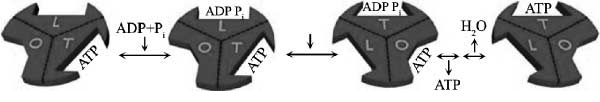
عملکرد موتور F1: بر اساس مکانیزم پیشنهاد شده برای این موتور، هر یک از زیرواحدهای β میتوانند در یکی از سه حالت O (باز)، L (شل) و T (بسته) قرار گیرند. زمانی که زیرواحد β در حالت O قرار دارد، موتور از لحاظ کاتالیستی، فعالیتی ندارد و تمایل بسیار کمی به اتصال با مولکولها (ADP و Pi) خواهد داشت. در حالت L، زیرواحد β پیوند ضعیفی با مولکولها برقرار میکند، اگرچههنوز از لحاظ کاتالیستی غیرفعال است. در حالت T، ملکول ADP و یون Pi به یکدیگر متصل میشوند و ملکول ATP را میسازند. در مرحله آخر، تغییر در شکل فضایی موتور، حالت T را به O تبدیل میکند و اجازه میدهد مولکول تولید شده ATP، از موتور جدا شود. شکل 3 مراحل مختلف این فرآیند را نشان میدهد.
موتور F0، عامل چرخش ستون γ است. چرخش ستون نانومتری γ که موتور F1 را به موتور F0 متصل کرده است، باعث تغییرشکل فضایی در موتور F1 میشود و نتیجه آن، تغییر حالت زیرواحدها است. همانطور که در شکل 2 ملاحظه میشود، شار یونی (و در برخی از موارد، شار یونی حاصل از یون هیدروژن) در طول غشا باعث چرخش موتور F0 میشود.
2-2- موتورهای میوزین و کینسین
در کنار موتورهای پروتئینی F0-F1-ATPase که به صورت چرخشی حرکت میکنند، موتورهای پروتئینی دیگری با حرکت خطی در طبیعت وجود دارند. این موتورها که انتقال اجزای مختلف مانند مواد غذایی، حرکت ماهیچهها، و تقسیم سلولی، از وظایف آنها به شمار میرود، برای حرکت خود نیاز به ملکول ATP دارند. این ماشینهای مولکولی کوچک به سه خانواده اصلی «کینسینها»، «میوزینها» و «داینینها» (dyneins) تقسیم بندی میشوند. در ادامه به توصیف دو خانواده اول پرداخته خواهد شد.
موتورهای میوزین: این موتورها خانوادهای گسترده از موتورهای پروتئینی هستند که قابلیت انتقال اجزای مختلف مانند مواد غذایی را در طول فیلامنت اکتینی (actin filament) دارند. فیلامنتهای اکتینی، لولههای ریز پروتئینی هستند که موتورهای مولکولی میوزین در طول آنها حرکت میکنند. این موتورها با هیدرولیز یا مصرف ملکول ATP، انرژی مورد نیاز برای حرکت خود را به دست میآورند. شمایی از تصویر موتور میوزین در شکل 4 نشان داده شده است. این موتوردارای دو سر (head) کروی مشابه است. هر یک از این دوسر که مستقل از یکدیگر عمل میکنند توسط یک بازوی اهرمی (level arm) از جنس پلیمر به یک رشته پلیمری متصل هستند. دو رشته پلیمری به صورت مارپیچ در هم پیچیدهاند و دو سر را متصل به یکدیگر نگه داشتهاند. دم موتور معمولاً به جزء منتقل شونده متصل است. لازم به ذکر است که در حین حرکت فقط یک سر موتور عمل میکند و سر دیگر آن آزادانه و بدون اتصال به فیلامنت اکتینی قرار دارد.

در شکل 5-(الف) موتور میوزین به همراه یک سر فعال آن (سبز رنگ) نشان داده شده است. این موتور بر روی فیلامنت اکتینی (آبی رنگ) حرکت میکند. بازوی اهرمی در این شکل به رنگ قرمز دیده میشود که سر موتور را به رشته مارپیچ سبز رنگ وصل کرده است. دلیل نام گذاری «بازو» این است که این بخش از موتور، قابلیت تغییر شکل فضایی دارد؛ هرچند به شکل بازو واقعی دیده نمیشود. بر اساس مکانیزم پیشنهاد شده، در ابتدا سر موتور با ATP واکنش داده و ADP و Pi را تولید میکند. در این زمان چون سر موتور به ADP متصل است، تمایلی به اتصال با فیلامنت اکتینی نازک ندارد. زمانی که موتور فعال به صورت تصادفی به فیلامنت اکتینی برخورد میکند، یون Pi بهجا مانده از واکنش آزاد میشود و اتصال برقرار میشود (شکل 5-الف-2). آزاد شدن یون فسفات باعث تغییرشکل فضایی بازوی اهرمی شده و آن را در جهت عکس ساعت میچرخاند. این چرخش، موتور مولکولی را به اندازه 10 نانومتر در طول فیلامنت اکتینی حرکت میدهد (شکل 5-الف-3). سپس مرکز کاتالیستی ADP خود را آزاد میکند و یک ATP جدید جای آن را میگیرد و باعث جدایش موتور از فیلامنت اکتینی نازک میشود (شکل 5-الف-4). بعد از آن، ATP به ADP و Pi تبدیل میشود و دوباره این پروسه تکرار میشود.
در صورتی که وظیفه موتور میوزین، حرکت ماهیچهها باشد، انتهای زنجیره پلیمری به جای اتصال به جزء منتقل شونده، به یک فیلامنت اکتینی نازک متصل است که به آن فیلامنت میوزین گفته میشود. این فیلامنت، موازی فیلامنت ثابت اکتینی است. در این صورت، حرکت موتور باعث لغزش فیلامنت میوزین نسبت به فیلامنت اکتینی ثابت میشود. نتیجه این لغزش، حرکت ماهیچه است.
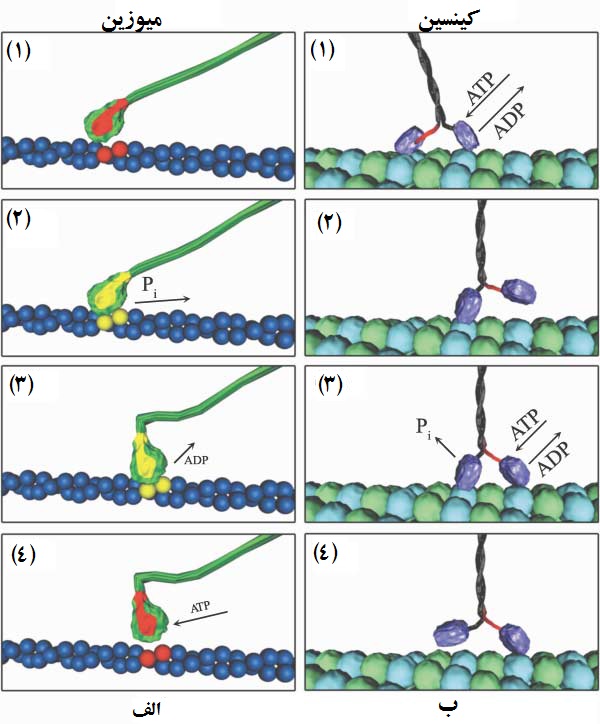
موتورهای کینسین: این موتورها با حرکت خطی در طول ریزلوله (microtubule) اجزای مختلف را جابهجا میکنند. ریزلولهها لولههایی به قطر 25 نانومتر هستند که از پروتئین خاصی به نام توبولین (tubulin) ساخته شدهاند. ریزلولهها قطبی هستند؛ یعنی یک سر منفی و یک سر مثبت دارند. کینسینها از سمت منفی به سمت مثبت حرکت میکنند. این موتورها نیز مانند میوزینها دارای دو سر هستند، با این تفاوت که هر دو سر در حرکت نقش دارند. شمایی از موتور کینسین در شکل 6 نشان داده شده است. هر کدام از دو سر موتور کینسین توسط اتصال گردنی به ساقه موتور وصل هستند. ذرات منتقلشونده نیز میتوانند به دم موتور وصل شوند. در حالت آزاد، هر دو سر موتور به مولکولهای جداگانه ADP وصل شدهاند و به صورت اتفاقی حرکت میکنند.

زمانی که یک سر موتور به ریزلوله برخورد میکند، به آن چسبیده و مولکول ADP خود را آزاد میکند. سپس یک مولکول ATP جدید، جای مولکول ADP را میگیرد (شکل 5-ب-1). این واکنش باعث چرخش اتصال گردنی میشود. اتصال گردنی با دور زدن موتور چسبیده به ریزلوله باعث چرخش و جلو آمدن سر دیگر موتور میشود (شکل 5-ب-2) و آن را به محل اتصال جلویی در طول ریزلوله متصل میکند. سپس سر اول موتور که به ATP متصل است، آن را هیدرولیز کرده، Pi حاصل از واکنش را جدا کرده، و ADP تولید شده را به خود متصل نگه میدارد. پس از آن، موتور جلویی ADP خود را با یک ATP جدید تعویض کرده (شکل 5-ب-3) و با چرخش اتصال گردنی باعث جلو آمدن سر عقبی میشود (شکل 5-ب-4). با تکرار این چرخه، موتور کینسین، با چرخش دو سر خود، روی ریزلوله قدم میزند. یک موتور کینسین قابلیت حرکت 100 قدم را بدون جدایش از ریز لوله دارد. همچنین سرعت حرکت این گونه موتورها میتواند به 1000 نانومتر بر ثانیه برسد. نیروی تولیدی نیز میتواند در حدود 5 تا 6 پیکو نیوتن باشد.
3- موتورهای مولکولی پایه DNA
ملکول DNA دو قابلیت مهم و کاربردی دارد: (1) شناخت مولکولی (molecular recognition)، و (2) خودآرایی. میتوان از همین قابلیتها برای ساخت موتورهای مولکولی استفاده کرد. نوکلئوتیدهای A و T و نیز C و G بر روی دو ss-DNA مختلف، تمایل دارند به یکدیگر متصل شوند. تنها زمانی یک ds-DNA پایدار و موثر تشکیل میشود که ترتیب نوکلئوتیدها بر روی هر دوشاخه ss-DNA به صورت مکمل (complementary) باشد؛ یعنی نوکلئوتیدهایی که در دو شاخه رو بروی هم قرار گرفتهاند، مکمل هم باشند (A روبروی T، و C روبروی G). بنابراین زمانی که مکمل ملکول ss-DNA در یک محلول باشد، نهایتاً یکدیگر را پیدا میکنند، به یکدیگر متصل میشوند و ملکول پایدار ds-DNA را تشکیل میدهند. از دیدگاه مکانیکی، اگر انرژی آزاد شده در اثر اتصال (یا زیپ شدن) دو ملکول مکمل DNA، برای بلند کردن یک بار فرضی به کار رود، میتواند نیرویی به اندازه حداکثر 15 پیکو نیوتن به وجود آورد. این مقدار نیرو در حد نیروهایی است که توسط موتورهای مولکولی پروتئینی تولید میشوند.
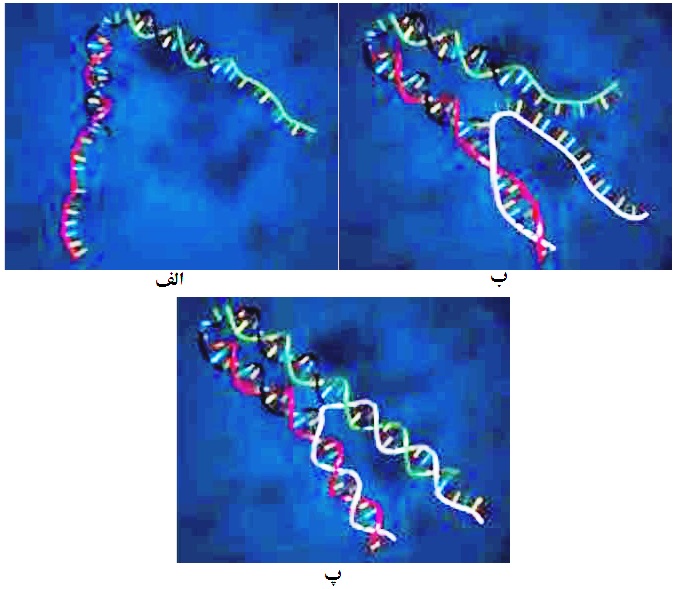
انبرک DNA (DNA Tweezers) یکی از موتورهای سنتز شده بر پایه DNA است که سوخت آن نیز ملکول DNA است. این موتور از سه رشته ss-DNA با نامهای A،B و C تشکیل شده است. رشتههای BوC به صورت موضعی بر روی رشته مرکزی A زیپ شدهاند (شکل 7-الف). سوخت این موتور، تک رشته ss-DNA به نام F است. طراحی این تکرشته به گونهای است که میتواند با بخشهای زیپ نشده B و C، زیپ شود (جفت شود). لذا زمانی که سوخت وارد محیطی میشود که موتور در آن قرار دارد (شکل 7-ب)، با بخشهای زیپ نشده B و C جفت میشود و موتور، شکل بسته به خود میگیرد (شکل 7-پ). در این زمان، اگر مکمل سوخت F، وارد محیط شود، سوخت را از موتور جدا کرده و موتور حالت باز (حالت اولیه) به خود میگیرد. این موتورها که قابلیت سوئیچ دارند، میتوانند در شناسایی انواع سوختها به کار روند؛ زیرا یک سوخت (ss-DNA) فقط برای یک نوع موتور مفید است. به عبارت دیگر، یک سوخت معین، فقط با یک موتور با طراحی مشخص، قابلیت جفت شدن دارد و میتواند آن را به کار بیاندازد.
4- موتورهای مولکولی غیرآلی (شیمیایی)
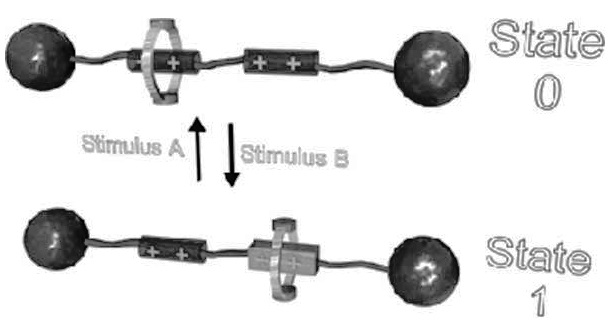
به عنوان مثال، موتور پایه روتاکسان (Rotaxanes)، یکی از موتورهای مولکولی سنتز شده است که قابلیت سوئیچ کردن دارند. شمایی از این موتور در شکل 8 نشان داده شده است. این دسته از موتورهای مولکولی ممولاً از دو بخش تشکیل شدهاند: (1) مولکول دمبلیشکل که دو گروه شیمیایی سنگین در دو انتهای آن است؛ و (2) مولکول سبک حلقویشکل که دور مولکول دمبلیشکل بین دو سر مولکول دمبلی قفل شده است. این مجموعه (ترکیب میله و دمبل) حاوی «دو مکان معین با جذب گزینشی» (selective) است و میتواند به حلقهها متصل شود. این اتصال میتواند از نوع قطبی و واندروالسی باشد. در حالت عادی، ترکیب حلقویشکل فقط به یکی از این دو مکان متصل است. با یک تحریک مناسب خارجی میتوان این اتصال را به گونهای تغییر داد که حلقه، برای حفظ پایداری ملکول، به مکان فعال دیگر جابجا شود. لذا با تحریک مناسب خارجی میتوان حلقه را بین دو مکان فعال جابجا کرد و منجر به سوئیچ موتور بین این دو حالت شد. تحریک مناسب خارجی میتواند ماهیت شیمیایی، الکتروشیمیایی و یا نوری داشته باشد. همچنین، نحوه اعمال تحریک میتواند روی سیکلها و سرعت موتور تاثیر بگذارد.
نتیجه گیری
موتورهای مولکولی انواع بسیار گستردهای دارند. تاکنون طیف وسیعی از موتورها در طبیعت کشف شده و بهخوبی مورد مطالعه قرار گرفتهاند. موتورهای مولکولی طبیعی بر پایه پروتئین هستند و سوخت اکثر آنها، ATP است. ATP منبع انرژی سیستمهای زنده است. یکی از راههای تولید این ماده، عملکرد ماشین مولکولی ATPase است. امروزه با شناخت موتورهای مولکولی طبیعی و تقلید از آنها، موتورهای مصنوعی بسیاری ساخته شدهاند؛ موتورهایی که میتوانند نیروهای مشابه یا حتی بیشتر از نیروهای تولیدی توسط گونههای طبیعی را تولید کنند. این موتورهای مصنوعی یا بر پایه DNA هستند و یا از نوع شیمیایی میباشند. سوخت اصلی موتورهای نوع اول، ملکولهای DNA است، اما موتورهای نوع دوم، انرژی خود را از طریق تحریک شیمیایی، الکتروشیمیایی، نور و یا تغییرات محیطی به دست میآورند.